Human CCL5/RANTES ELISA Kit
 Protocol Booklet
Protocol Booklet
Key features and details
- Specification:
- Sensitivity:
- Standard Curve Range:
- Standard Curve Gradient:
- Number of Incubations:
- Sample Volume:
- Assay type:
- Operation Duration:
-
Brand:

CAT.NO. : AEH0027
RMB Please choose
RMB Please choose
Size:
Trail, Bulk size or Custom requests Please contact us
*产品价格可能会有所调整,请以品牌方官网实时更新的价格为准,以确保准确性。
 Product Details
Product Details
Background
RANTES (Regulated upon Activation, Normal T cell Expressed and presumably Secreted), also known as CCL5, is a member of the "CC" subfamily of chemokines. It plays a primary role in the inflammatory immune response via its ability to chemoattract leukocytes and modulate their function. The cDNA for RANTES was initially discovered by subtractive hybridization as a T cell specific sequence. Human RANTES cDNA encodes a highly basic 91 amino acid (aa) residue precursor polypeptide with a 23 aa hydrophobic signal peptide that is cleaved to generate the 68 aa mature protein. Human RANTES exhibits approximately 85% homology with mouse RANTES at the deduced aa level.
RANTES is a potent chemoattractant for a number of different cell types including unstimulated CD4+/CD45RO+ memory T cells and stimulated CD4+ and CD8+ T cells with naive and memory phenotypes, NK cells, basophils, eosinophils, dendritic cells, mast cells, monocytes, and microglia. In addition to its effects on migration, RANTES can activate a number of cell types including T cells, monocytes, neutrophils, NK cells, dendritic cells, and astrocytes. T cell activation generally requires relatively high RANTES concentrations (~ 1 μM) and is dependent upon aggregation of the molecule and association with cell surface glycosaminoglycans (GAGs). Whether this[missing some words, probably 'activity occurs in vivo'] remains unclear although in mice, intraperitoneally injected RANTES mutants that are unable to aggregate and/or bind GAG, are not capable of attracting leukocytes when compared to wild-type controls. Other in vivo studies show that RANTES knockout mice exhibit deficient recruitment of leukocytes to sites of acute inflammation. - RANTES, is known to interact with four identified seven transmembrane G-protein coupled receptors: CCR1, CCR3, CCR4, and CCR5 (22-25). RANTES stimulation can initiate a variety of signaling cascades that are cell context dependent. For instance, in T-cells, RANTES can stimulate elevations of intracellular Ca2+, and activation of focal adhesion kinase (FAK), protein kinase A, PI3-kinase, Rho GTPase, and JAK/STAT signaling pathways. The cytomegalovirus protein US28 exhibits significant homology with CC chemokine receptors and is capable of binding RANTES. Membrane-spanning US28 can, depending on the context, signal in a constitutive manner, bind RANTES and initiate G-protein-mediated signaling cascades, or sequester RANTES and potentially alter inflammatory responses. -
The RANTES receptor CCR5 is also the primary co-receptor for R5 (M-tropic) variants of HIV-1. It has been demonstrated that RANTES, as well as the other CCR5 ligands, macrophage inflammatory protein (MIP)-1 alpha and MIP-1 beta, can competitively inhibit CCR5/HIV-1 interaction and suppress viral infection in vitro. These effects apparently do not require fully intact signaling from the CCR5 receptor. Consequently, modified forms of RANTES and non-peptide compounds that block the interaction of HIV-1 with CCR5 show promise for future therapies. In contrast, several reports show that RANTES can enhance in vitro replication of X4 (T-tropic) variants of HIV-1 that use CXCR4 as a co-receptor rather than CCR5. This activity usually requires relatively high RANTES concentrations (~μM) and is dependent upon interaction with cell surface GAGs, oligomerization, and activation of tyrosine kinase and MAP kinase signaling cascades.
RANTES is a potent chemoattractant for a number of different cell types including unstimulated CD4+/CD45RO+ memory T cells and stimulated CD4+ and CD8+ T cells with naive and memory phenotypes, NK cells, basophils, eosinophils, dendritic cells, mast cells, monocytes, and microglia. In addition to its effects on migration, RANTES can activate a number of cell types including T cells, monocytes, neutrophils, NK cells, dendritic cells, and astrocytes. T cell activation generally requires relatively high RANTES concentrations (~ 1 μM) and is dependent upon aggregation of the molecule and association with cell surface glycosaminoglycans (GAGs). Whether this[missing some words, probably 'activity occurs in vivo'] remains unclear although in mice, intraperitoneally injected RANTES mutants that are unable to aggregate and/or bind GAG, are not capable of attracting leukocytes when compared to wild-type controls. Other in vivo studies show that RANTES knockout mice exhibit deficient recruitment of leukocytes to sites of acute inflammation. - RANTES, is known to interact with four identified seven transmembrane G-protein coupled receptors: CCR1, CCR3, CCR4, and CCR5 (22-25). RANTES stimulation can initiate a variety of signaling cascades that are cell context dependent. For instance, in T-cells, RANTES can stimulate elevations of intracellular Ca2+, and activation of focal adhesion kinase (FAK), protein kinase A, PI3-kinase, Rho GTPase, and JAK/STAT signaling pathways. The cytomegalovirus protein US28 exhibits significant homology with CC chemokine receptors and is capable of binding RANTES. Membrane-spanning US28 can, depending on the context, signal in a constitutive manner, bind RANTES and initiate G-protein-mediated signaling cascades, or sequester RANTES and potentially alter inflammatory responses. -
The RANTES receptor CCR5 is also the primary co-receptor for R5 (M-tropic) variants of HIV-1. It has been demonstrated that RANTES, as well as the other CCR5 ligands, macrophage inflammatory protein (MIP)-1 alpha and MIP-1 beta, can competitively inhibit CCR5/HIV-1 interaction and suppress viral infection in vitro. These effects apparently do not require fully intact signaling from the CCR5 receptor. Consequently, modified forms of RANTES and non-peptide compounds that block the interaction of HIV-1 with CCR5 show promise for future therapies. In contrast, several reports show that RANTES can enhance in vitro replication of X4 (T-tropic) variants of HIV-1 that use CXCR4 as a co-receptor rather than CCR5. This activity usually requires relatively high RANTES concentrations (~μM) and is dependent upon interaction with cell surface GAGs, oligomerization, and activation of tyrosine kinase and MAP kinase signaling cascades.
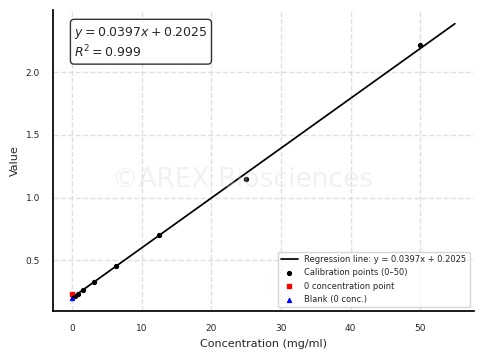
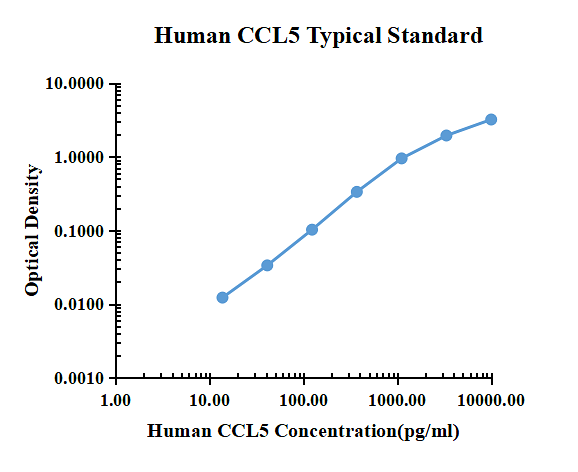
Typical data
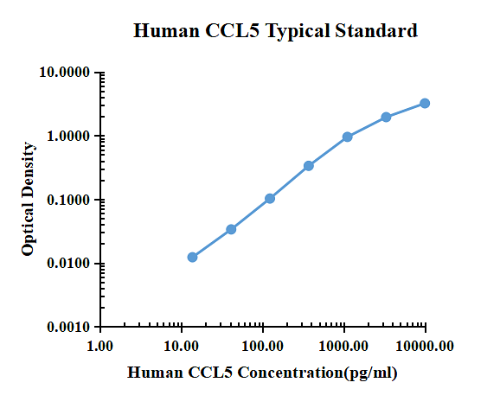
| pg/ml | O.D. | Average | Corrected | |
| 0.00 | 0.0151 | 0.0162 | 0.0157 | |
| 13.72 | 0.0271 | 0.0288 | 0.0280 | 0.0123 |
| 41.15 | 0.0489 | 0.0496 | 0.0493 | 0.0336 |
| 123.46 | 0.1175 | 0.1189 | 0.1182 | 0.1025 |
| 370.37 | 0.3486 | 0.3520 | 0.3503 | 0.3346 |
| 1111.11 | 1.0108 | 0.9250 | 0.9679 | 0.9522 |
| 3333.33 | 1.8759 | 2.0533 | 1.9646 | 1.9489 |
| 10000.00 | 3.2985 | 3.1585 | 3.2285 | 3.2128 |
Precision
| Intra-assay Precision | Inter-assay Precision | |||||
| Sample Number | S1 | S2 | S3 | S1 | S2 | S3 |
| 22 | 22 | 22 | 6 | 6 | 6 | |
| Average(pg/ml) | 426.6 | 1631.6 | 3396 | 702.2 | 3412 | 8842.1 |
| Standard Deviation | 37.6 | 128.2 | 288.6 | 44.6 | 194.4 | 506.5 |
| Coefficient of Variation(%) | 8.8 | 7.9 | 8.5 | 6.4 | 5.7 | 5.7 |
Intra-assay Precision (Precision within an assay) Three samples of known concentration were tested twenty times on one plate to assess intra-assay precision.
Inter-assay Precision (Precision between assays) Three samples of known concentration were tested six times on one plate to assess intra-assay precision.
Spike Recovery
The spike recovery was evaluated by spiking 3 levels of human CCL5 into health human serum sample. The un-spiked serum was used as blank in this experiment.
The recovery ranged from 90% to 106% with an overall mean recovery of 97%.
The recovery ranged from 90% to 106% with an overall mean recovery of 97%.
Sample Values
| Sample Matrix | Sample Evaluated | Range (pg/ml) | Detectable (%) | Mean of Detectable (pg/ml) |
| Serum | 30 | 943.0-38560.8 | 100 | 21207.7 |
Serum/Plasma – Thirty samples from apparently healthy volunteers were evaluated for the presence of CCL5 in this assay. No medical histories were available for the donors.
New Products